Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].

Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).

AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
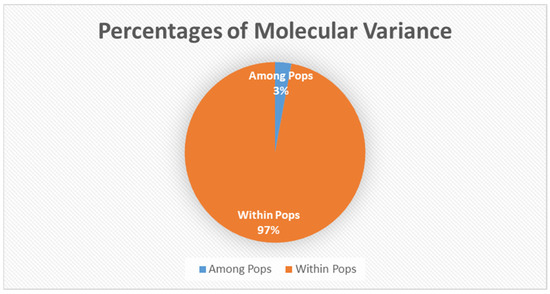
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
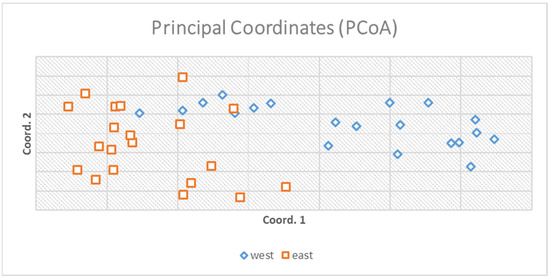
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]
Чужеродные и инвазивные растения угрожают целостности природных экосистем, поскольку большинство из них являются инвазивными, конкурируют с местными растениями и во многих случаях занимают их место в экосистеме. Инвазивные виды характеризуются быстрым распространением за счет производства большого количества потомства, которое может рассеиваться на большие расстояния [ 1 ]. Более того, они имеют высокую выживаемость благодаря своей пластичности, способной реагировать на различные условия окружающей среды. Кроме того, некоторые природные явления могут способствовать их распространению, например, наводнения, ветер, поверхностная эрозия и т. д.
На распространение инвазивных видов также существенно повлияла деятельность человека [ 2 ]. С древних времен чужеродные растения случайно или намеренно переносились в новые места обитания и культивировались как садовые или декоративные растения [ 3 ]. Их распространению способствует также деятельность человека, роль которой двоякая: транспортировка размножающегося материала и изменение среды обитания. Главное преимущество заключается в том, что у них нет физических врагов, которые бы контролировали их население, чтобы они в полной мере использовали свой конкурентный потенциал. Некоторые инвазивные растения также развивают новые механизмы взаимодействия с природными экосистемами [ 4 ]. Одним из таких механизмов является продукция аллелопатических веществ и их выброс в экосистему [ 5].
Amorpha L. ( Fabaceae Juss.: Amorpheae Britains .) — род из 16 видов, произрастающих в Северной Америке [ 6 ]. Эти растения могут представлять собой кустарники, полукустарники или травянистые многолетние растения и встречаются в различных средах обитания, включая прибрежные коридоры, саванны и луга. Многие виды Amorpha имеют очень ограниченное распространение, а некоторые считаются находящимися под угрозой исчезновения [ 7 ].
Amorpha fruticosa L. (куст индиго) — многолетний листопадный кустарник, принадлежащий к семейству Fabaceae и произрастающий в юго-западной части Северной Америки [ 8 ]. Куст индиго был завезен в Европу как декоративное растение [ 5 ]. Его используют как лекарство для лечения ожогов и экземы [ 9 ]. В последние годы ему уделяется особое внимание из-за его биологической активности. Исследования показали, что это растение богато полифенолами и содержит биологически активные ингредиенты. Он также обладает противораковой, противовоспалительной и противомикробной активностью [ 10] .]. Куст индиго – растение, имеющее большую декоративную ценность и используемое при обустройстве городских ландшафтов и при охране склонов. Он может выжить в сухих почвах, но предпочитает берега рек и опушки затопленных лесов, где легко распространяется водой, поскольку его семенные коробочки плавучие [11 ] . Его высокая толерантность к различным климатическим условиям, а также отличная способность к размножению дают ему возможность с легкостью расти за пределами своего естественного распространения [ 12 ]. Сегодня он признан одним из наиболее инвазивных видов в Европе, оказывающим серьезное воздействие на местные экосистемы [ 11 ]. Это светолюбивое растение, конкурирующее за свет, образующее густые заросли, затмевающие местную флору, а это приводит к снижению биоразнообразия [ 11, 12 , 13 ]. Из-за ущерба, который он наносит природным экосистемам и водному хозяйству, следует немедленно принять меры по сокращению/контролю его популяции.
В Греции трансграничные реки, такие как Нестос, Аксиос и др., являются транспортными коридорами для чужеродных видов растений, которые расширяют свой ареал за счет воды [ 14 ]. Однако, помимо конкретного вида транспорта, в последние годы в Грецию были интродуцированы многие зарубежные виды растений, как намеренно (например, декоративные и лекарственные растения), так и непреднамеренно, используя различные способы интродукции [2 ] . Хотя число чужеродных видов велико, не все из них являются инвазивными. Из 343 чужеродных видов только 50 проявляют инвазионное поведение, и большинство из них имеют американское происхождение [ 15 ].
Нестос — трансграничная река Болгарии и Северной Греции. Берет начало от горы Рила на юге Болгарии и проходит между горными хребтами Аймос и Родопы. Его длина составляет 234 км, из них 130 км находится на территории Греции [ 16 ]. Прежде чем впасть в море, он простирается через равнину Хрисуполис и расширяется как дельтовая система с пресноводными озерами, образующими дельту Нестоса. В этой зоне существует приоритетная среда обитания 91E0 (Приложение I Директивы 92/43/EEC), в которой представлены два подтипа: подтип ольховых насаждений ( Alnus Glutinosa ) и подтип тополя белого ( Populus alba ) и вяза ( Ulmusminor ).) стоит. Первый подтип состоит из чистых насаждений ольхи, произрастающих на легких и умеренно легких почвах с очень хорошими физическими свойствами, которые можно охарактеризовать как Calcaric fluvisols. При этом уровень грунтовых вод относительно высокий (90–95 см). Этот подтип встречается между дамбами, местами вблизи берегов рек или в местах, удаленных от них на несколько десятков метров. Второй подтип составляет значительную часть тугайного леса и располагается за пределами дамб. Он состоит из насаждений белого тополя и вяза, которые растут на глубоких и умеренно легких почвах с хорошими физическими свойствами, принадлежащих к типу почв Eutric Fluvisol, без проблем с щелочностью и засоленностью. Уровень грунтовых вод низкий, но сильно колеблется [ 17 ].
Территория вдоль реки существенно изменилась со времен Второй мировой войны [ 18 ]. В 1945 году на этой территории преобладали леса и водно-болотные угодья. В 1960-е годы площадь сельскохозяйственных угодий увеличилась, что привело к уменьшению лесных площадей. Также произошло значительное сокращение площади водно-болотных угодий из-за их преобразования в пастбища. Однако, несмотря на сильное упадок природной экосистемы, в этом районе по-прежнему сохраняется самый обширный речной лес в Греции. Кроме того, равнина реки Нестос имеет важное экологическое значение из-за множества важных мест обитания. Он был классифицирован как водно-болотное угодье международного значения и включен в Рамсарскую конвенцию. Он также защищен Бернской конвенцией, директивами Европейской комиссии и законодательством Греции. Кроме того, он принадлежит к сети Natura 2000.19 ].
В Греции Amorpha fruticosa L. считается инвазивным видом, поскольку это растение способно распространяться на обширной территории. В настоящей работе изучено генетическое разнообразие популяций Amorpha fruticosa L., произрастающих вдоль реки Нестос. Наши цели заключались в том, чтобы (1) определить уровень генетической дифференциации, если она существует, между двумя берегами реки: западным и восточным; (2) оценить степень инбридинга в каждой популяции, а также эффективный размер популяции; и (3) обсудить последствия расширения конкретных видов в естественной экосистеме и, более конкретно, в популяциях местных видов в этом районе.
2. Материалы и методы
2.1. Растительный материал
Сбор образцов листьев проведен на 20 особях Amorpha fruticosa из 2 популяций района реки Нестос (ориентировочные координаты западной популяции A: X: 565671, Y: 4527927 и восточной популяции: X: 566918, Y: 4525481). Экстракцию ДНК из свежей хвои проводили с помощью набора Macherey Nagel Plant (Düren, Северный Рейн-Вестфалия, Германия) в соответствии с протоколом производителя.
2.2. Процедура ПФЛП
Всего 200 нг геномной ДНК фрагментировали расщеплением 4 ед. ферментов EcoRI и MseI при температуре 37°С в течение 3 часов. Расщепленные фрагменты ДНК затем объединяли с адаптерами EcoRI и MseI и полученную смесь лигировали с использованием ДНК-лигазы Т4 от New England Biolabs (Ипсвич, Массачусетс, США). Этот этап лигирования проводили при температуре 26°C в течение 3 часов. Полученную в результате этого процесса ДНК использовали в качестве первичной матрицы для анализа AFLP.
Для этапа преамплификации использовали пару праймеров на основе последовательностей адаптеров EcoRI и MseI с дополнительным селективным нуклеотидом на 3'-конце (EcoRI+A и MseI+C). ПЦР с предварительной амплификацией проводили в общем объеме 20 мкл, который содержал 1X буфер Kapa Taq, 0,2 мМ каждого dNTP, 2,5 мМ MgCl 2., 30 нг каждого праймера (EcoRI+A и MseI+C), 1 единица ДНК-полимеразы Taq от Kapa Biosystems (расположена в Уилмингтоне, Массачусетс, США) и 5 мкл разведенных фрагментов ДНК, полученных в результате реакции расщепления и лигирования. . Условия циклирования ПЦР были следующими: начальная денатурация при 95 °С в течение 30 с, затем 32 цикла денатурации при 95 °С в течение 30 с, отжиг при 56 °С в течение 30 с, удлинение при 72 °С в течение 1 мин. и заключительный этап удлинения при 72 ° C в течение 10 минут. Для подтверждения амплификации образец реакции объемом 5 мкл подвергали электрофорезу в агарозном геле, а оставшиеся 15 мкл разбавляли в 5 раз ТЕ-буфером.
Селективную амплификацию проводили в реакционных объемах объемом 10 мкл, состоящих из 3 мкл разбавленной преселективной матрицы, и с использованием тех же условий реакции, что и на стадии преселективной амплификации, за исключением использования 30 нг праймера MseI и 5 нг праймера EcoRI. за реакцию. Программа циклической селективной амплификации осуществлялась с использованием термоциклера BioRad (Hercules, Калифорния, США) по следующей схеме: начальный цикл денатурации при 95 °C в течение 30 с, отжиг при 65 °C в течение 30 с и удлинение при 95 °C в течение 30 с. 72 °С в течение 1 мин. За этим следовали двенадцать циклов денатурации при 95°С в течение 30 с, отжиг, начинающийся при 65°С в течение 30 с и уменьшающийся на 0,75°С в каждом цикле, и удлинение при 72°С в течение 1 мин. Наконец, двадцать три цикла денатурации при 95°С в течение 30 с, отжиг при 56°С в течение 30 с,
Смеси продуктов f-AFLP подвергали денатурации путем обработки их формамидом при температуре 94 °С в течение 2 мин. Впоследствии эти денатурированные смеси разделяли электрофорезом с использованием генетического анализатора ABI Prism 3730xl, который производится Applied Biosystems (Уолтем, Массачусетс, США). Использовали десять комбинаций селективных праймеров AFLP. Затем каждый отдельный гибрид исследовали для определения наличия или отсутствия конкретных фрагментов. Размер обнаруженных фрагментов определяли с использованием программы GeneMapper 4.0 вместе с внутренним стандартом, известным как GS 500 LIZ, который также был предоставлен Applied Biosystems (Уолтем, Массачусетс, США). Чтобы свести к минимуму влияние потенциальной гомоплазии размеров,20 ].
2.3. Индекс регенерации
Присутствие и плотность A. fruticosa оценивались в приоритетном местообитании 91E0* в районе Нестос на северо-востоке Греции ( рис. 1 ). Пятнадцать (15) участков, каждый площадью 706,86 м 2 , были выбраны случайным образом и равномерно размещены по всей территории обитания 91E0*. В дальнейшем был проведен анализ состава и структуры древесных пород на этих участках. A. fruticosa присутствовала на четырех участках из 15, из которых три располагались на западном берегу реки и один на восточном. У особей A. fruticosa измеряли высоту дерева (м), плотность (особей га -1 ) и индекс регенерации (IR) . IR рассчитывали путем умножения плотности A. fruticosa (на м2 ) по средней высоте A. fruticosa (см) и позволяет оценить степень регенерации [ 20 ].
Рисунок 1. Amorpha fruticosa , распространенная на приоритетном местообитании 91Ε0* в дельте Нестоса, на северо-востоке Греции.
2.4. Сбор данных и статистический анализ данных AFLP
Для преобразования информации о размере аллелей, полученной с помощью GeneMapper 4.0 (Applied Biosystems, США), в двоичный формат использовался макрос AFLP Excel. Этот макрос присваивал значение «1» присутствующим фрагментам и «0» отсутствующим. Чтобы свести к минимуму влияние потенциальной гомоплазии размеров, подсчитывали и подвергали дополнительному изучению только согласованные фрагменты, попадающие в диапазон от 150 до 500 пар оснований [ 21 ]. Кроме того, GenAlex v6.0 [ 22 ] использовался для расчета таких параметров, как процент полиморфных полос (P), информационный индекс Шеннона (I) и генетическое разнообразие (He). GenAlex v6.0 также использовался для проведения анализа молекулярной дисперсии (AMOVA), а также для расчета генетических расстояний и анализа главных координат (PCoA).
Подробно, анализ главных координат (PCoA) представляет собой многомерный метод, который позволяет находить и отображать основные закономерности в многомерном наборе данных (например, множественных локусах и множественных образцах). Математика сложна, но, по сути, PCoA — это процесс, с помощью которого основные оси вариаций располагаются в многомерном наборе данных. Для многомерных наборов данных каждая последующая ось объясняет пропорционально меньшую часть общей вариации, так что при наличии отдельных групп первые две или три оси обычно выявляют большую часть разделения между ними. Предусмотрено четыре различных варианта: два основаны на преобразовании матрицы расстояний в ковариационную матрицу, а два работают непосредственно на основе входной матрицы расстояний. Матрица расстояний используется как пул для расчета распределения по осям [22 ].
2.5. Информация о среде обитания 91E0*
Местообитание 91E0* в дельте реки Нестос простирается на небольших высотах (<100 м над уровнем моря) с поверхностями от умеренного до плоского рельефа (уклон 0–10%) (выборка личных данных Коракаки Э. и Карецоса Г.). В его состав входят доминирующие виды деревьев, такие как Fraxinus excelsior L. и Alnus Glutinosa (L.) Gaertn., а также Fraxinus angustifolia Vahl, Salix alba L., Populus alba L., Pinus nigra JF Arnold, Juglans regia L., Cornus sanguinea. L., Ulmus major Mill. и Quercus robur subsp. pedunculiflora (К. Кох) Меницкого. Существует множество вьющихся растений (например,Periploca graeca L. и др.), а травянистый ярус неизменно включает множество видов (например, Aegopodium podagraria L., Hedera helix L., Lysimachia punctata L., Circaea lutetiana L., Arum maculatum L. и др.) (Solomou Выборка персональных данных А. и Карецоса Г.). Дельта Нестоса образована аллювиальными отложениями реки Нестос. Речные отложения состоят из аллювиальных отложений глин, песков и гравия, тогда как пойменные отложения состоят преимущественно из глин и песков [ 23 ].
3. Результаты
3.1. Генетические результаты
Семь комбинаций праймеров AFLP дали 313 локусов для двух популяций. По результатам AFLP, восточная популяция имела полиморфизм 83,39%, а западная — 83,07%. Средняя ожидаемая гетерозиготность (He) составляла 0,127, а западная популяция имела среднюю гетерозиготность немного выше (0,136), чем восточная (0,119). Параметры генетического разнообразия представлены в Таблице 1 вместе с эпигенетическими результатами.
Таблица 1. Среднее значение и стандартная ошибка (SE) для Na (количество аллелей), Ne (количество эффективных аллелей), I (информационный индекс Шеннона) и He (ожидаемая гетерозиготность).
AMOVA (анализ молекулярной дисперсии) определил 97% генетических вариаций внутри популяций и 3% только между популяциями ( рис. 2 ). Анализ основных координат объяснил 16,89% дисперсии ( рис. 3 ).
Рисунок 2. Анализ молекулярной дисперсии среди популяций и внутри них: дисперсия среди популяций составила 3%, а внутри - 97%. Генетическая вариация фрагментов AFLP, использованных для анализа AMOVA.
Рисунок 3. Анализ основных координат объяснил 16,89% общей изменчивости, и были сформулированы две небольшие группы в зависимости от происхождения популяций. На рисунке обозначена небольшая группировка двух популяций, полученная на основе матрицы генетических расстояний генотипов.
3.2. Плотность и индекс регенерации
На четырех участках было зарегистрировано в общей сложности 233 особи A. fruticosa , в результате чего средняя плотность составила 824,07 растений на гектар. Примечательно, что плотность A. fruticosa и индекс регенерации (IR) оказались выше у популяции, проживающей на западном берегу реки, по сравнению с восточным берегом реки. Однако стоит отметить, что средняя высота деревьев была выше на восточном берегу реки, как указано в Таблице 2 . В аллювиальном лесу наиболее широко распространено возобновление чужеродных видов A. fruticosa и A. negundo (личное сообщение Коракаки).
Таблица 2. Средняя высота, средняя плотность и средний индекс регенерации (IR).
4. Дискуссия
Сообщается, что ложное индиго ( Amorpha fruticosa L.) является неместным [ 15 ], и недавние наблюдения зафиксировали его присутствие на значительных территориях вдоль рек Ардас, Эврос и Нестос на северо-востоке Греции, что указывает на его завоз из соседней Болгарии. , возможно, через речной транспорт генетического материала. В прибрежных лесах реки Нестос на северо-востоке Греции A. fruticosa демонстрирует значительную динамику регенерации и, по-видимому, подавляет растительность в важных приоритетных местообитаниях.
В этом исследовании мы оценили генетическое разнообразие двух популяций с западного и восточного берегов реки Нестос, чтобы оценить, имеют ли они различия в генетических параметрах или имеют ли они схожие индексы генетического разнообразия. По оценкам локусов AFLP, гетерозиготность западной популяции была немного выше, но существенно не отличалась от восточной популяции. Более того, как показал анализ AMOVA, большая часть генетических вариаций наблюдалась внутри популяций (97%). Анализ основных координат объяснил лишь 16,89% общей изменчивости, и были сформулированы две небольшие группы по происхождению популяций. Главный вывод генетического анализа заключался в том, что западные и восточные популяции имеют общий генетический фонд, который поступает с водой через реку Нестос.
Что касается аналогичных исследований, авторы не обнаружили никаких упоминаний о генетическом разнообразии с использованием молекулярных маркеров растений A. fruticosa . Однако есть одно исследование [ 24 ], в котором они измерили 47 агрономических и морфологических признаков энергии биомассы. Результаты показали, что на все признаки повлияли образцы, и это ключ, который также может демонстрировать генетическое содержание, которое, вероятно, влияет на изменчивость. Кроме того, были проведены исследования экспрессии генов, связанных с засухой [ 25 ], фенольных компонентов [ 26 ], протеомного анализа симбиотических белков [ 27 ] и исследований рака [ 28 , 29] .]. Вышеупомянутые исследования с одной точки зрения указывают на значение A. fruticosa , хотя мы не можем исключить негативное воздействие, которое он оказывает на экосистемы из-за его быстрого распространения. Этот инвазивный вид может проникнуть в луговые сообщества и быстро превратить среду обитания в густые и однородные заросли. Он вытесняет местные растения и нарушает естественную сукцессию растений и биоразнообразие. Это представляет собой серьезную угрозу, поскольку подавляет местные виды деревьев в приоритетном аллювиальном лесу дельты Нестоса (91E0*). Кроме того, это ставит под угрозу выживание видов Emys orbcularis , Testudo Hermanni и Callimorpha Quadripunctaria *, поскольку они зависят от этих мест обитания.
Согласно нашим выводам, A. fruticosa более активно развивается на западном берегу реки Нестос, что представляет значительный риск для биоразнообразия и подавления местных видов деревьев в аллювиальных лесах региона Нестос. Таким образом, любые природоохранные меры и действия по управлению лесами, направленные на ограничение распространения A. fruticosa , должны быть приоритетными на этом берегу реки. Меры по сохранению необходимы для защиты и поддержания этой естественной среды обитания, которая представляет значительный интерес для сообщества. Действия также должны быть направлены на повышение осведомленности об угрозах, с которыми сталкиваются аллювиальные леса, и на обеспечение их долгосрочной защиты и сохранения. Этот результат ИК дает нам первое указание на то, что A. fruticosaбудет более обширным на западной стороне, и результат небольшой, но более высокой гетерозиготности согласуется с текущим результатом. С 2021 года Институт средиземноморских лесных экосистем (ELGO-DIMITRA) разрабатывает различные экспериментальные методы лечения, чтобы изучить возможные решения по ограничению распространения A. fruticosa .
Наконец, в этом исследовании мы обнаружили аналогичный генетический пул с восточного и западного берегов реки Нестос, который, вероятно, происходит из Болгарии. Будущие исследования должны также включать генетический анализ этих болгарских популяций. Считается необходимым продолжить исследования и изучение A. fruticosa в ближайшие годы, чтобы было более полное знание этого вида и его влияния на экосистему, чтобы затем можно было создать и создать систему управления этим видом. его преимущества можно использовать, не оказывая негативного воздействия на экосистему.
5. Выводы
Первое сообщение о генетическом разнообразии и распространении чужеродного вида Amorpha fruticosa L. было впервые использовано за рекой Нестос в Греции. Мы изучили две популяции, одну с западной стороны и одну с восточной стороны, с маркерами AFLP, и не обнаружили дифференциации между ними. Это указывает на то, что, вероятно, один генетический пул был передан через реку из Болгарии, и будущие исследования должны также охватывать болгарские популяции. Кроме того, проведенная первая оценка состояния регенерации также указывает на расширение вида. Это обязательный шаг для оценки и предложения мер управления с целью сохранения и ограничения распространения вида, который оказывает существенное негативное воздействие на среду обитания 91E0*.
Вклад автора
Концептуализация, EVA и EK; методология, EVA, EK, EM, ADS, GM и ГК; программное обеспечение, EVA, EM и EK; проверка, все; ресурсы, EVA и EK; курирование данных, EVA и EK; письмо — подготовка оригинального проекта, EVA, EK, EM и ADS; написание — просмотр и редактирование, все; надзор, EVA и EK; администрация проекта, ЭК; приобретение финансирования, Е.К. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось проектом LIFE PRIMED, LIFE17 NAT/GR/000511, который софинансируется Греческим зеленым фондом (GRFU).
Заявление о доступности данных
Данные недоступны.
Конфликт интересов
Авторы объявили, что нет никаких конфликтов интересов.
Рекомендации
- Ричардсон, DM; Пышек, П.; Рейманек, М.; Барбур, Миннесота; Панетта, Флорида; Уэст, К.Дж. Натурализация и вторжение чужеродных растений: концепции и определения. Дайверы. Распредел. 2000 , 6 , 93–107. [ Академия Google ] [ CrossRef ]
- Коракаки, Э.; Легакис, А.; Кацаневакис, С.; Кулелис, П.; Аврамиду, Э.; Сулиоти, Н.; Петракис, П. Инвазивные чужеродные виды Греции. Обсервация инвазивных чужеродных видов. Проблемы вокруг света 2021 , 3 , 124–189. [ Академика Google ]
- Мияваки, С.; Вашитани, И. Инвазивные чужеродные виды растений в прибрежных районах Японии: вклад сельскохозяйственных сорняков, видов восстановления растительности и аквакультурных видов. Глоб. Окружающая среда. Рез.-англ. Эд. 2004 , 8 , 89–101. [ Академика Google ]
- Каллауэй, РМ; Ашехуг, Э.Т. Инвазивные растения против их новых и старых соседей: механизм экзотического вторжения. Наука 2000 , 290 , 521–523. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Иванеску, Б.; Лунгу, К.; Власе, Л.; Градинару, AC; Тухилус, К. ВЭЖХ-анализ фенольных соединений, антиоксидантной и противомикробной активности Amorpha fruticosa L. Экстракты. Дж. Плант Дев. 2019 , 26 , 77. [ Академия Google ] [ CrossRef ]
- Штрауб, Южная Каролина; Извините, бакалавр; Уикли, А.С. Amorpha confusa , новое название старой Amorpha (Fabaceae: Amorpheae). Дж. Бот. Рез. Инст. Техас, 2009 г. , 151–155. [ Академика Google ]
- Штрауб, Южная Каролина; Дойл, Дж. Дж. Молекулярная филогенетика Amorpha (Fabaceae): оценка монофилии, видовых отношений и полиплоидного происхождения. Мол. Филогенетика Эвол. 2014 , 76 , 49–66. [ Академия Google ] [ CrossRef ]
- Лю, Дж.; Ван, ET; Chen, WX Разнообразные ризобии, связанные с древесными бобовыми культурами Wisteria sinensis , Cercis Racemosa и Amorpha fruticosa, выращиваемыми в умеренной зоне Китая. Сист. Прил. Микробиол. 2005 , 28 , 465–477. [ Академия Google ] [ CrossRef ]
- Китайский, П. Словарь китайской Materia Medica ; Китайская научно-техническая медицинская пресса: Пекин, Китай, 1997. [ Google Scholar ]
- Митчер, Луизиана; Парк, Ю.Х.; Альшамма, А.; Хадсон, ПБ; Хаас, Т. Аморфрутин a и b, бибензиловые противомикробные средства из Amorpha fruticosa . Фитохимия 1981 , 20 , 781–785. [ Академия Google ] [ CrossRef ]
- Крпан, АП; Томашич, Ж.; Башич Палкович, П. Биопотенциал индигобуша ( Amorpha fruticosa L.) – второй год исследования. Список Шумарского. 2011 , 135 , 103–112. [ Академика Google ]
- Кожухарова Е.; Матковский, А.; Возняк, Д.; Симеонова Р.; Найчов З.; Малайнер, К.; Мокан, А.; Набави, С.М.; Атанасов А.Г. Amorpha fruticosa – вредоносное инвазионное чужеродное растение в Европе или лекарственное растение от болезней обмена веществ? Передний. Фармакол. 2017 , 8 , 333. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Сигетвари, К. Первые шаги по восстановлению пойменного луга после десятилетия доминирования инвазивного кустарника-трансформера Amorpha fruticosa L. Tiscia 2002 , 33 , 67–77. [ Академика Google ]
- Педашенко, Л.П.; Апостолова, И.И.; Васильев К.В. Невселенность Amorpha fruticosa в различных местообитаниях нижнего Дуная. Фитол. Балк. 2012 , 18 , 285–291. [ Академика Google ]
- Ариануцу, М.; Базос, И.; Делипетру, П.; Коккорис, Ю. Инопланетная флора Греции: таксономия, особенности жизни и предпочтения среды обитания. Биол. Вторжения 2010 , 12 , 3525–3549. [ Академия Google ] [ CrossRef ]
- Самарас, АГ; Коутитас, К.Г. Моделирование воздействия на прибрежную морфологию управления водными ресурсами в трансграничных речных бассейнах: пример реки Нестос. Менеджер. Окружающая среда. Квал. Межд. Дж. 2008 , 19 , 455–466. [ Академия Google ] [ CrossRef ]
- Декамп, Х.; Декамп, О. Средиземноморские прибрежные леса ; Сохранение водно-болотных угодий Средиземноморья; Тур дю Валат: Арль, Франция, 2001 г. [ Google Scholar ]
- Маллинис, Г.; Эмманолудис, Д.; Яннакопулос, В.; Марис, Ф.; Куциас, Н. Картирование и интерпретация исторических изменений растительного покрова/землепользования на участке «Натура 2000» с использованием данных наблюдений за Землей: пример дельты Нестоса, Греция. Прил. геогр. 2011 , 31 , 312–320. [ Академия Google ] [ CrossRef ]
- Дафис, С.; Папастергиаду, Э.; Георгиу, К.; Бабалонас, Д.; Георгиадис, Т.; Папагеоргиу, М.; Лазариду, Т.; Циаусси, В. Греческий проект «среды обитания» Natura 2000: Обзор. Пожизненный контракт B4-32000/94/756 ; Комиссия Европейских Сообществ, Музей естественной истории Гуландриса, Центр греческих биотопов и водно-болотных угодий; Springer: Берлин/Гейдельберг, Германия, 1996. [ Google Scholar ]
- Бьянки, Л.; Пачи, М. Эволюционная динамика и тосканское обучение: Sintesi di quarant'anni di richerche. Для.-Ж. Сильвик. Для. Экол. 2008 , 5 , 122. [ Google Scholar ] [ CrossRef ]
- Векеманс, X.; Боуэнс, Т.; Лемэр, М.; Ролдан-Руис, И. Данные маркеров амплифицированного полиморфизма длин фрагментов (AFLP) указывают на гомоплазию по размеру и взаимосвязь между степенью гомоплазии и размером фрагмента. Мол. Экол. 2002 , 11 , 139–151. [ Академия Google ] [ CrossRef ]
- Пикалл, РОД; Смаус, ЧП Genalex 6: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований. Мол. Экол. Примечания 2006 , 6 , 288–295. [ Академия Google ] [ CrossRef ]
- Тафтсоглу, М.; Валканиотис, С.; Папатанасиу, Г.; Климис, Н.; Докас, И. Подробная карта подверженности разжижению дельты реки Нестос, Фракия, Греция, основанная на поверхностной геологии и геоморфологии. Geosciences 2022 , 12 , 361. [ Академия Google ] [ CrossRef ]
- ДеХаан, Л.; Эльке, Н.; Шиффер, К.; Вайз, Д.; ДеХаан, Р. Оценка разнообразия североамериканских образцов ложного индиго ( Amorpha fruticosa L.) для корма и биомассы. Жене. Ресурс. Кроп Эвол. 2006 , 53 , 1463–1476. [ Академия Google ] [ CrossRef ]
- Ли, М.; Лю, З.; Лю, К.; Чжу, Ф.; Ван, К.; Ван, З.; Ли, Х.; Лан, X.; Гуан, К. Засухоустойчивость табака, сверхэкспрессирующего ген afnac1 Amorpha fruticosa Linn. Передний. Наука о растениях. 2022 , 13 , 980171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Дат, Северная Каролина; Ли, Дж.-Х.; Лук-порей.; Хонг, Ю.-С.; Ким, Ю.Х.; Ли, Дж. Дж. Фенольные компоненты Amorpha fruticosa , которые ингибируют активацию nf-κb и связанную с ней экспрессию генов. Дж. Нэт. Прод. 2008 , 71 , 1696–1700. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Песня, Ф.; Ци, Д.; Лю, X.; Конг, X.; Гао, Ю.; Чжоу, З.; Ву, К. Протеомный анализ симбиотических белков Glomus mosseae и Amorpha fruticosa . наук. Rep. 2015 , 5 , 18031. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Тодорова Т.; Бояджиев, К.; Шкондров А.; Пырванова П.; Димитрова, М.; Ионкова И.; Крастева И.; Кожухарова Е.; Чанкова, С. Скрининг экстрактов Amorpha fruticosa и Ailanthus altissima на генотоксичность/антигенотоксичность, мутагенность/антимутагенность и канцерогенность/антиканцерогенность. БиоРиск 2022 , 17 , 201–212. [ Академия Google ] [ CrossRef ]
- Цветкович, Д.М.; Йованкич, СП; Милутинович, М.Г.; Никодиевич, Д.Д.; Грбович, Ф.Дж.; Чирич, АР; Топузович, доктор медицинских наук; Маркович, С.Д. Противоинвазивная активность Robinia pseudoacacia L. и Amorpha fruticosa L. На клеточной линии рака молочной железы mda-mb-231. Биология 2019 , 74 , 915–928. [ Академия Google ] [ CrossRef ]