
- Категория: Блог
- Просмотров: 590
Оптический микроскоп - это удивительное устройство, которое позволяет увидеть мир невидимый невооруженным глазом. С помощью оптического микроскопа можно рассмотреть мельчайшие детали объектов, которые невозможно увидеть обычным способом.
Этот инструмент играет важную роль в сферах науки, медицины и промышленности. Оптический микроскоп позволяет исследователям изучать клетки, микроорганизмы, минералы и другие объекты меньше размером чем ваши волосы. Благодаря микроскопу мы можем расширить свои знания о мире и обогатить нашу жизнь новыми открытиями.
На сайте Радоника вы найдете широкий выбор оптических микроскопов различных производителей и моделей. У нас вы сможете найти микроскопы для начинающих и опытных исследователей, а также специализированные микроскопы для медицинских и научных целей.

Мы предлагаем только качественные товары, которые прошли проверку и имеют высокие технические характеристики. Наши оптические микроскопы обеспечат вам четкое изображение и увеличение, что поможет вам сделать точные исследования.
Приобретая оптический микроскоп на нашем сайте, вы получите не только качественный продукт, но и профессиональную консультацию и быструю доставку. Мы гарантируем удовлетворение вашего запроса и отличное обслуживание.
Исследуйте мир с оптическим микроскопом от Радоника. Потратьте время на изучение невидимых деталей и откройте для себя удивительный мир микрокосмоса.
Преимущества микроскопов компании Радоника
1. Высокое качество изготовления: микроскопы от компании Радоника производятся с использованием самых современных технологий и материалов, что обеспечивает высокую точность и надежность работы.
2. Широкий выбор моделей: компания Радоника предлагает широкий ассортимент микроскопов различных типов и назначений, что позволяет выбрать оптимальное решение для конкретных задач и потребностей пользователя.
3. Превосходное оптическое качество: микроскопы от Радоника оснащены высококачественной оптикой, что обеспечивает четкое и детальное изображение обследуемого объекта.
4. Простота в использовании: микроскопы компании Радоника удобны в эксплуатации и обладают интуитивно понятным интерфейсом, что делает работу с ними максимально удобной и эффективной.
5. Надежное обслуживание и поддержка: компания Радоника предоставляет гарантийное и постгарантийное обслуживание микроскопов, а также консультационную поддержку для пользователей.
- Категория: Блог
- Просмотров: 584
Анализатор удельной поверхности – это прибор или программное обеспечение, которое позволяет определить удельную поверхность материала. Удельная поверхность – это показатель, характеризующий площадь поверхности материала в единицу массы. Он используется в различных областях, таких как химия, физика, материаловедение и биология.
Анализатор удельной поверхности может работать на основе различных принципов измерения, таких как измерение площади поверхности методом Брэтчера-Эммета-Теллера (BET), измерение углового наклона капиллярной структуры поверхности, измерение контактного угла жидкости на поверхности и другие.
Использование анализатора удельной поверхности позволяет получить информацию о взаимодействии материала с окружающей средой, его адсорбционных и адсорбционно-десорбционных свойствах, а также о его микроструктуре.
Таким образом, анализатор удельной поверхности является важным инструментом для исследования и контроля свойств материалов, что позволяет повысить их качество и оптимизировать их использование в различных областях науки и техники.
Сорбция пористых материалов - это процесс, при котором одно вещество (сорбат) поглощается и задерживается внутри пористой структуры другого вещества (сорбента). Пористые материалы, такие как активированный уголь, глины, молекулярные ситечатые материалы и другие, обладают высокой поверхностной площадью и могут эффективно сорбировать различные вещества из газовой или жидкой фазы.
Процесс сорбции зависит от многих параметров, включая размер и форму пор, химический состав сорбента и сорбата, температуру, давление и концентрацию сорбата. Сорбция может использоваться для очистки воды, воздуха, пищевых продуктов, фармацевтических препаратов и других материалов от различных загрязнителей и вредных веществ.
Метод BET (Brunauer-Emmett-Teller) - это техника адсорбции газов, используемая для изучения поверхностных свойств материалов, таких как размер пор и площадь поверхности.
Анализ размеров пор методом BET происходит следующим образом:
1. Сначала материал подвергают адсорбции газа при различных относительных давлениях.
2. Затем измеряется объем газа, адсорбированного материалом, и строится изотерма адсорбции.
3. По данным изотермы рассчитывают площадь поверхности материала и объем пор.
4. Расчеты проводятся с использованием уравнения Брунера-Эммета-Теллера, которое учитывает молекулярные взаимодействия между газом и поверхностью материала.
Изучение размеров пор методом BET является полезным для определения структуры материала, его поверхностных свойств и взаимодействия с окружающей средой. Полученные данные могут использоваться в различных областях, таких как катализ, фармацевтика, пищевая промышленность и другие.
- Категория: Блог
- Просмотров: 529
Анализаторы металлов EXPLORER 5000 от компании Радоника - это современное и надежное оборудование, которое позволяет с точностью определить состав металлических материалов. Они широко применяются в различных отраслях промышленности, где требуется быстрый и точный анализ металлов и сплавов.
Одним из лидирующих производителей анализаторов металлов является компания "Analyser", которая предлагает широкий выбор оборудования для анализа металлов различных типов и структур. Используя современные технологии и методики, анализаторы "Analyser" обеспечивают высокую точность и надежность результатов.
Преимущества использования анализаторов металлов:
1. Быстрота и удобство. Анализаторы металлов позволяют провести анализ состава материала за считанные минуты, что значительно экономит время и упрощает процесс контроля качества.
2. Точность и надежность. Современные анализаторы обеспечивают высокую точность результатов анализа, что позволяет избежать ошибок и недочетов при определении состава металлических материалов.
3. Многофункциональность. Анализаторы металлов могут использоваться для анализа различных типов металлов, сплавов и структур, что делает их универсальным и эффективным инструментом для проведения контроля качества.
Если вам необходимо приобрести анализатор металлов, обратитесь в компанию "Analyser", где вам помогут подобрать подходящее оборудование и обеспечат качественный сервис и поддержку.
Оптимизируйте работу вашего предприятия с помощью современного оборудования для анализа металлов от ведущего производителя. Убедитесь в надежности и качестве анализаторов металлов и повысьте эффективность производственных процессов!
- Категория: Блог
- Просмотров: 505
Перистальтические насосы – это устройства, которые работают на принципе перистальтики, то есть сжатия и расширения гибкой трубки, через которую проходит жидкость. При сжатии трубки создается давление, которое выталкивает жидкость вперед, а при расширении трубки создается вакуум, благодаря чему жидкость подается в насос. Перистальтические насосы используются в различных отраслях промышленности, медицине, лабораторных и исследовательских целях, а также в системах оборудования для водных ресурсов. Они отличаются от других типов насосов своей способностью перекачивать агрессивные, вязкие, осажденные или чувствительные ксорбенты жидкости без их загрязнения или повреждения.
Перистальтические насосы – это современное оборудование, которое широко используется в различных отраслях промышленности и медицине. Они отличаются высокой точностью дозирования, надежностью и долговечностью, что делает их незаменимым инструментом для передачи жидкостей и газов.
Одним из ключевых преимуществ перистальтических насосов является их способность работать без контакта с перекачиваемым материалом. Это позволяет избежать загрязнения насоса и предотвращает загрязнение жидкости или газа. Кроме того, они обеспечивают плавную и равномерную подачу материала, что делает их идеальным выбором для задач, где требуется точное дозирование.
Перистальтические насосы часто применяются в таких отраслях, как фармацевтика, пищевая промышленность, лабораторные исследования, водоочистка, медицина и другие. Они могут быть использованы для передачи различных жидкостей, включая агрессивные и вязкие среды, благодаря своей уникальной конструкции и химической стойкости.
Если вам требуется надежное и эффективное оборудование для передачи жидкостей и газов, обратитесь к специалистам в области перистальтических насосов. Они помогут вам выбрать оптимальное решение для ваших потребностей и обеспечат качественное обслуживание и поддержку.
Перистальтический насос работает на основе принципа перистальтики, который подразумевает передачу жидкости с помощью сжатия и расширения гибкой трубки или шланга. Внутри этой трубки находится жидность, которая перемещается вперед благодаря перистальтическим движениям, создаваемым специальным ротором или валиком.
Процесс работы перистальтического насоса можно разделить на несколько этапов:
1. В начале цикла на входе насоса трубка расширяется под действием вакуума, создаваемого сдвигом ротора или валика.
2. Затем ротор или валик начинает сжимать трубку по всей длине, что приводит к перемещению жидкости вперед.
3. После этого трубка снова расширяется, обеспечивая прочистку и наполнение ее новой порцией жидкости.
4. Процесс сжатия и расширения трубки повторяется циклически, обеспечивая постоянное перемещение жидкости в нужном направлении.
Перистальтические насосы обеспечивают точное и безопасное перемещение жидкостей, не контактируя с ними напрямую и предотвращая возможные загрязнения или перекачку. Это делает их идеальным выбором для широкого спектра применений, включая медицинскую и лабораторную технику, пищевую и фармацевтическую промышленность, а также для подачи реагентов и других химических веществ.
Не откладывайте на будущее – обновите свою технику и повысьте производительность вашего производства с помощью перистальтических насосов прямо сейчас!
- Категория: Блог
- Просмотров: 784
1. Введение
Пространственно-временная оценка уязвимости подземных вод
2. Методология
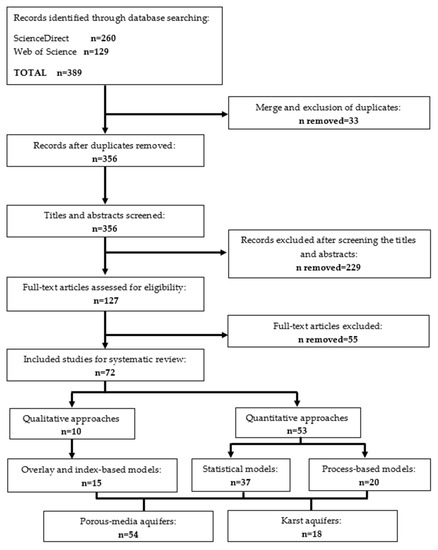
3. Результаты и их обсуждение
3.1. Оценка уязвимости пористого водоносного горизонта
3.1.1. Модели наложения и индексы
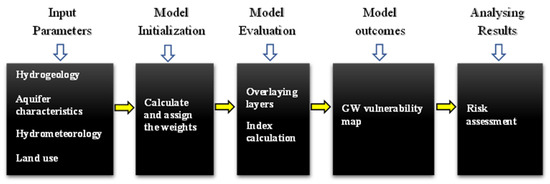

3.1.2. Статистические модели
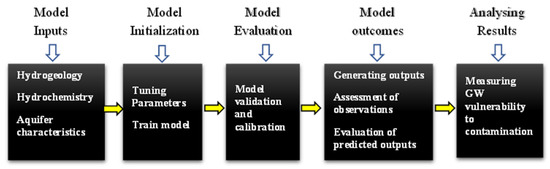
Пространственные интерполяции
Многомерная статистика
Регрессионные модели
Искусственный интеллект
Пространственная автокорреляция
Байесовские сети
Другие статистические методы
Физические/процессные методы
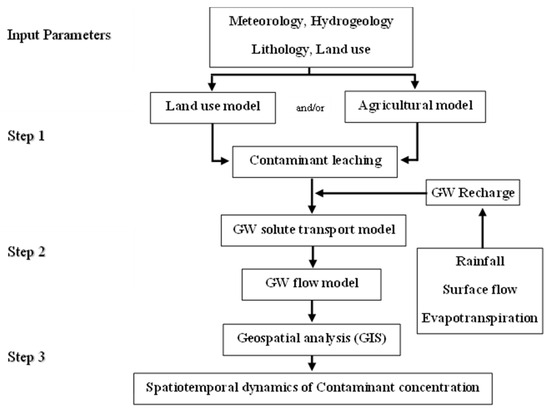
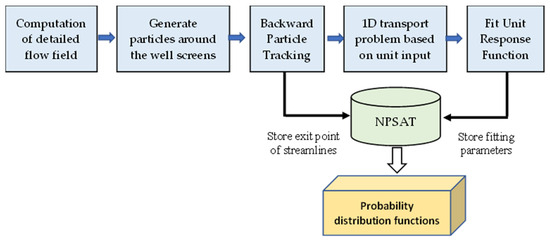
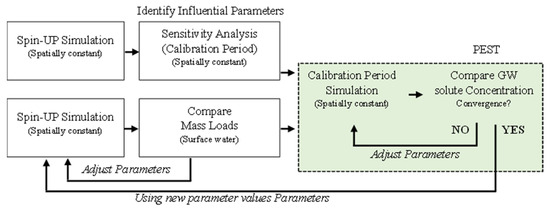
3.2. Оценка уязвимости карстового водоносного горизонта
3.2.1. Метод наложения и индексный метод
3.2.2. Статистический метод
3.2.3. Процессный метод
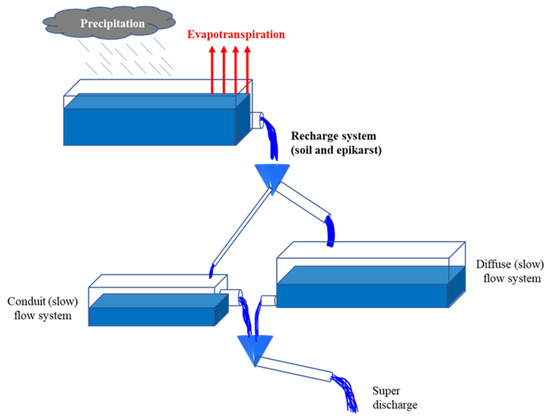
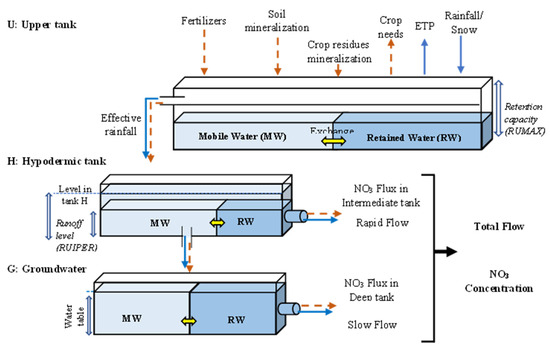
4. Обзор методов динамической оценки уязвимости подземных вод
5. Ограничения и будущие направления
6. Выводы
Дополнительные материалы
Вклад автора
Финансирование
Благодарности
Конфликт интересов
Рекомендации
- Лалл, У.; Жоссет, Л.; Руссо, Т. Краткий обзор мировых проблем подземных вод. Анну. Преподобный Окружающий. Ресурс. 2020 , 45 , 171–194. [ Академия Google ] [ CrossRef ]
- ИГРАЦ. Обзор подземных вод: невидимое становится видимым. 2018. Доступно онлайн: https://www.un-igrac.org/sites/default/files/resources/files/Groundwater%20overview%20-%20Making%20the%20invisible%20visible_Print.pdf (по состоянию на 29 июля 2023 г.) .
- Денадхаялан, Н.; Лин, КК; Салех, Т.А. Последние достижения в области функционализированных углеродных точек в направлении разработки эффективных материалов для приложений зондирования и катализа. Малый 2020 , 16 , e1905767. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Мачивал, Д.; Джа, МК; Сингх, вице-президент; Мохан, К. Оценка и картирование уязвимости подземных вод к загрязнению: Текущее состояние и проблемы. Науки о Земле. Ред. 2018 г. , 185 , 901–927. [ Академия Google ] [ CrossRef ]
- Мачивал, Д.; Клотье, В.; Гюлер, К.; Казакис, Н. Обзор интегрированных с ГИС статистических методов оценки и защиты качества подземных вод. Окружающая среда. Науки о Земле. 2018 , 77 , 681. [ Google Scholar ] [ CrossRef ]
- Равбар, Н.; Гольдшайдер, Н. Предлагаемая методология картирования уязвимости и риска загрязнения для защиты карстовых водоносных горизонтов в Словении (Предлог методологии картирания ранливости в veganja za onesnaženje voda za varovanje kraških vodonosnikov v Sloveniji). Acta Carsologica 2007 , 36 , 397–411. [ Академика Google ]
- Стивенацци, С.; Масетти, М.; Нгием, СВ; Соричетта, А. Карты уязвимости земных опорных устройств в зависимости от метода, зависящего от времени использования спутниковых скаттерометров. Гидрогеол. Дж. 2015 , 23 , 631–647. [ Академия Google ] [ CrossRef ]
- Бутчер, К.; Хуггенбергер, П. Внутренняя оценка уязвимости карстовых территорий: подход к численному моделированию. Водный ресурс. Рез. 2008 , 44 . [ Академия Google ] [ CrossRef ]
- Бутчер, К.; Хуггенбергер, П. Расширенная оценка уязвимости карстовых территорий путем сочетания подходов картирования и моделирования. наук. Тотальная среда. 2009 , 407 , 1153–1163. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Маргат, Дж.; ван дер Гун, Дж. Подземные воды во всем мире: географический обзор ; CRC Press: Бока-Ратон, Флорида, США, 2013. [ Google Scholar ]
- Феттер, CW «Прикладная гидрогеология» , 4-е изд.; Прентис Холл: Хобокен, Нью-Джерси, США, 2001 г. [ Google Scholar ]
- Гош, Северная Каролина Геогенное загрязнение и технологии безопасного снабжения питьевой водой BT — Вода и санитария в новом тысячелетии ; Нат, К.Дж., Шарма, вице-президент, ред.; Спрингер: Нью-Дели, Индия, 2017 г.; стр. 81–95. [ Академия Google ] [ CrossRef ]
- Блумфилд, Япония; Уильямс, Р.Дж.; Гудди, округ Колумбия; Кейп, JN; Гуха, П. Влияние изменения климата на судьбу и поведение пестицидов в поверхностных и грунтовых водах – взгляд Великобритании. наук. Тотальная среда. 2006 , 369 , 163–177. [ Академия Google ] [ CrossRef ]
- Ковалевский В.С. Влияние изменений климата на подземные воды. Водный ресурс. 2007 , 34 , 140–152. [ Академия Google ] [ CrossRef ]
- Аурели, А.; Танигучи, М. Оценка ресурсов подземных вод под воздействием человечества и изменений климата. Париж. 2006. Доступно онлайн: https://iwlearn.net/resolveuid/01d776e51f524e6fa99c85b17686aece (по состоянию на 29 июля 2023 г.).
- Грин, Т.Р. Связь изменения климата и подземных вод. В «Интегрированном управлении подземными водами: концепции, подходы и проблемы» ; Джейкман А.Дж., Баррето О., Хант Р.Дж., Ринаудо Дж.Д., Росс А., ред.; Springer: Берлин/Гейдельберг, Германия, 2016 г.; стр. 97–141. [ Академия Google ] [ CrossRef ]
- Стигтер, Тайвань; Рибейро, Л.; Дилл, AMMC Оценка внутреннего и специфического метода оценки уязвимости в сравнении с уровнями засоления грунтовых вод и уровнями загрязнения нитратами в двух сельскохозяйственных регионах на юге Португалии. Гидрогеол. Дж. 2006 , 14 , 79–99. [ Академия Google ] [ CrossRef ]
- Маргат, Дж. Vulnerabilite des Nappes d'eau Souterraine a la Pollution (Уязвимость подземных вод к загрязнению) ; Bureau de Recherches Géologiques et Minères (BRGM): Орлеан, Франция, 1968. [ Google Scholar ]
- Врба, Дж.; Запорозец А. Руководство по картированию уязвимости подземных вод ; Х. Хайзе: Ганновер, Германия, 1994. [ Google Scholar ]
- Национальный исследовательский совет. Оценка уязвимости подземных вод: прогнозирование относительного потенциала загрязнения в условиях неопределенности ; The National Academies Press: Вашингтон, округ Колумбия, США, 1993. [ Google Scholar ] [ CrossRef ]
- Бутчер, К.; Хуггенбергер, П. Моделирование временной изменчивости уязвимости карстовых подземных вод с последствиями для изменения климата. Окружающая среда. наук. Технол. 2009 , 43 , 1665–1669. [ Академия Google ] [ CrossRef ]
- Дерфлигер, Н.; Жаннен, П.-Ю.; Звален, Ф. Оценка уязвимости воды в карстовых средах: новый метод определения защитных зон с использованием многоатрибутного подхода и инструментов ГИС (метод EPIK) ; Springer: Берлин/Гейдельберг, Германия, 1999. [ Google Scholar ]
- Гогу, РЦ; Дассарг, А. Текущие тенденции и будущие проблемы в оценке уязвимости подземных вод с использованием методов наложения и индексов ; Springer: Berlin/Heidelberg, Germany, 2000. [ Google Scholar ]
- Андерсон, член парламента; Весснер, В.В. (ред.) Глава 1 – Введение. В прикладном моделировании подземных вод ; Академическое издательство: Сан-Диего, Калифорния, США, 2002 г.; стр. 1–11. [ Академия Google ] [ CrossRef ]
- Буфекан, А.; Сайги О. Применение методов наложения и индексирования уязвимости подземных вод на равнине Джиджель (Алжир). Подземные воды 2018 , 56 , 143–156. [ Академия Google ] [ CrossRef ]
- Масетти, М.; Стерлаккини, С.; Баллабио, К.; Сорикетта, А.; Поли, С. Влияние порогового значения при использовании статистических методов для оценки уязвимости подземных вод. наук. Тотальная среда. 2009 , 407 , 3836–3846. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Хаан, Статистические методы КТ в гидрологии ; Издательство Университета штата Айова: Эймс, Айова, США, 1977. [ Google Scholar ]
- Соторникова Р.; Врба, Дж. Концепция карт уязвимостей. В материалах Международной конференции по уязвимости почвы и подземных вод к загрязнителям, Гаага, Нидерланды, 30 марта – 3 апреля 1987 г.; стр. 471–476. [ Академика Google ]
- Барбулеску, А. Оценка уязвимости подземных вод: КРАСТИЧНЫЕ и КРАСТИЧНЫЕ методы: обзор. Вода 2020 , 12 , 1356. [ Академика Google ] [ CrossRef ]
- Хагбин, М.; Шарафати, А.; Диксон, Б.; Кумар, В. Применение мягких вычислительных моделей для моделирования загрязнения нитратами подземных вод: комплексный обзор, оценка и будущие возможности. Арх. Вычислить. Методы англ. 2020 , 28 , 3569–3591. [ Академия Google ] [ CrossRef ]
- Либерати, А.; Альтман, Д.Г.; Тецлафф, Дж.; Малроу, К.; Гётше, ПК; Иоаннидис, JPA; Кларк, М.; Деверо, Пи Джей; Клейнен, Дж.; Мохер, Д. Заявление PRISMA о систематических обзорах и метаанализах исследований, оценивающих медицинские вмешательства: объяснение и разработка. Анна. Стажер. Мед. 2009 , 62 , Е1–Е34. [ Академия Google ] [ CrossRef ]
- Нан, З.; Бо, Л.; Чанглай, X. Пространственно-временные изменения загрязнения подземных вод с использованием IEA-UEF в городских районах города Цзилинь, северо-восточный Китай. Наука о воде. Технол. Водоснабжение 2016 , 16 , 1277–1286. [ Академия Google ] [ CrossRef ]
- Махмуд, К.; Уль-Хак, З.; Батул, ЮАР; Рана, AD; Тарик, С. Применение временной ГИС для отслеживания областей, вызывающих серьезную обеспокоенность в отношении загрязнения подземных вод. Окружающая среда. Earth Sci 2016 , 75 , 33. [ Google Scholar ] [ CrossRef ]
- Чатурведи, А.; Бхаттачарджи, С.; Сингх, АК; Кумар, В. Новый подход к индексированию загрязнения подземных вод тяжелыми металлами. Экол. Индик. 2018 , 87 , 323–331. [ Академия Google ] [ CrossRef ]
- Миси, А.; Гуминдога, В.; Хоко, З. Оценка потенциала и уязвимости подземных вод в водосборном бассейне Верхнего Маньяме в Зимбабве. Физ. хим. Земля 2018 , 105 , 72–83. [ Академия Google ] [ CrossRef ]
- Дуттагупта, С.; Мукерджи, А.; Дас, К.; Дутта, А.; Бхаттачарья, А.; Бхаттачарья, Дж. Оценка уязвимости подземных вод к загрязнению пестицидами в аллювиальном водоносном горизонте бассейна Западной Бенгалии, Индия, с использованием метода наложения и индекса. Геохимия 2020 , 80 , 125601. [ Google Scholar ] [ CrosRef ]
- Ву, Т.Д.; Ни, КФ; Ли, туалет; Труонг, М.Х. Модифицированный метод наложения индексов для оценки пространственно-временных изменений уязвимости подземных вод и риска загрязнения подземных вод в районах с различной сельскохозяйственной деятельностью. Вода 2019 , 11 , 2492. [ Академия Google ] [ CrossRef ]
- Нистор М.М. Уязвимость ресурсов подземных вод при изменении климата Паннонского бассейна. Гео-Спат. Инф. наук. 2019 , 22 , 345–358. [ Академия Google ] [ CrossRef ]
- Хайду, И.; Нистор, М.М. Оценка уязвимости подземных вод в регионе Гранд-Эст, Франция. Кват. Межд. 2020 , 547 , 86–100. [ Академия Google ] [ CrossRef ]
- Бахрами, М.; Зарей, Арканзас; Ростами, Ф. Временная и пространственная оценка загрязнения подземных вод нитратами с помощью индекса загрязнения нитратами (NPI) и ГИС (пример: равнина Фасаруд, южный Иран). Конв. Geochem Health 2020 , 42 , 3119–3130. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Мендес, член парламента; Рибейру, Л. Картирование вероятности нитратов в аллювиальной системе северного водоносного горизонта реки Тежу (Португалия) с использованием дизъюнктивного кригинга. наук. Тотальная среда. 2010 , 408 , 1021–1034. [ Академия Google ] [ CrossRef ]
- Курунц, А.; Эршахин, С.; Сонмез, Северная Каролина; Каман, Х.; Уз, И.; Уз, BY; Аслан, Г.Е. Сезонные изменения пространственных изменений некоторых переменных качества подземных вод в большом орошаемом прибрежном средиземноморском регионе Турции. наук. Тотальная среда. 2016 , 554–555 , 53–63. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Ди Гуардо, А.; Финицио, А. Подход мони-моделирования для управления риском выщелачивания пестицидов подземными водами в региональном масштабе. наук. Тотальная среда. 2016 , 545–546 , 200–209. [ Академия Google ] [ CrossRef ]
- Ди Гуардо, А.; Финицио, А. Клиент-серверное программное обеспечение для выявления уязвимости подземных вод к пестицидам на региональном уровне. наук. Тотальная среда. 2015 , 530–531 , 247–256. [ Академия Google ] [ CrossRef ]
- Чицазан, М.; Табари, MMR; Эйлбейги, М. Анализ временных и пространственных изменений содержания нитратов в грунтовых водах и развитие шлейфа их загрязнения: тематическое исследование в водоносном горизонте Карадж. Окружающая среда. Науки о Земле. 2017 , 76 , 391. [ Google Scholar ] [ CrossRef ]
- Нааз, А.; Кумар, Б.; Нараян, К.; Шукла, К.; Аншумали. Оценка загрязнения фторидами подземных вод аридных и полуаридных районов тоналит-тронджемитового ряда Центральной Индии. Экспо Здоровье 2015 , 7 , 545–556. [ Академия Google ] [ CrossRef ]
- Он, Х.; Цянь, Дж.; Лю, З.; Лу, Ю.; Ма, Л.; Чжао, В.; Канг, Б. Временно-пространственная эволюция загрязнения подземных вод азотом за семь лет в высокоурбанизированном городе на юге Китая. Бык. Окружающая среда. Контам. Токсикол. 2017 , 99 , 753–759. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Ричардс, Г.; Гилмор, штат Техас; Миттельштет, Арканзас; Мессер, ТЛ; Сноу, Д.Д. Динамика нитратов в базовом стоке внутри водоразделов сельскохозяйственного ручья в Небраске, США. Сельское хозяйство. Экосист. Окружающая среда. 2021 , 308 , 107223. [ Академия Google ] [ CrossRef ]
- Арслан, Х.; Туран, Н.А.; Демир, Ю.; Гюнгор, А.; Цемек, Б. Оценка пространственных и сезонных изменений загрязнения подземных вод нитратами сельскохозяйственных земель с помощью обычных и индикаторных методов кригинга. Арх. Агрон. Почвоведение. 2017 , 63 , 907–917. [ Академия Google ] [ CrossRef ]
- Гош, М.; Пал, ДК; Сантра, С.К. Пространственное картирование и моделирование загрязнения подземных вод мышьяком и оценка риска с помощью метода геопространственной интерполяции. Окружающая среда. Дев. Поддерживать. 2020 , 22 , 2861–2880. [ Академия Google ] [ CrossRef ]
- Менхен, А.; Лас Эрас, JD; Алдай, Дж. Дж. Загрязнение пестицидами подземных вод в пилотном бассейне Европейского Союза на реке Хукар (ЮВ-Испания). Окружающая среда. Monit Assess 2017 , 189 , 146. [ Google Scholar ] [ CrossRef ]
- Чен, С.; Ву, В.; Ху, К.; Ли, В. Влияние изменений в землепользовании и ресурсах ирригационной воды на загрязнение нитратами неглубоких грунтовых вод в масштабе округа. Экол. Сложный. 2010 , 7 , 131–138. [ Академия Google ] [ CrossRef ]
- Бой-Рура, М.; Менсио, А.; Мас-Пла, Дж. Временной анализ данных родниковой воды для оценки поступления нитратов в грунтовые воды в сельскохозяйственных районах (Осона, северо-восток Испании). наук. Тотальная среда. 2013 , 452–453 , 433–445. [ Академия Google ] [ CrossRef ]
- Ли, К.; Чжан, Х.; Го, С.; Фу, К.; Ляо, Л.; Сюй, Ю.; Ченг, С. Распределение источников загрязнения подземных вод с использованием анализа главных компонентов на территории с разнообразным землепользованием на юго-западе Китая. Окружающая среда. наук. Загрязнение. Рез. 2020 , 27 , 9000–9011. [ Академия Google ] [ CrossRef ]
- Аззеллино, А.; Коломбо, Л.; Ломби, С.; Маркези, В.; Пиана, А.; Андреа, М.; Альберти, Л. Диффузное загрязнение подземных вод в функциональных городских районах: необходимость определения фоновых уровней антропогенного диффузного загрязнения. наук. Тотальная среда. 2019 , 656 , 1207–1222. [ Академия Google ] [ CrossRef ]
- Мастрочикко, М.; Ди Джузеппе, Д.; Винченци, Ф.; Коломбани, Н.; Кастальделли, Г. Происхождение и судьба хлората в неглубоких грунтовых водах под сельскохозяйственными ландшафтами. Окружающая среда. Загрязнение. 2017 , 231 , 1453–1462. [ Академия Google ] [ CrossRef ]
- Юнг, Х.В.; Юн, СТ; Ким, К.Х.; О, СС; Канг, К.Г. Роль непроницаемого слоя в контроле химического состава подземных вод в базальтовом водоносном горизонте под сельскохозяйственным полем, остров Чеджу, Южная Корея. Прил. Геохим. 2014 , 45 , 82–93. [ Академия Google ] [ CrossRef ]
- Мачивал, Д.; Джа, М.К. Выявление источников загрязнения подземных вод в системе водоносных горизонтов твердых пород с использованием многомерного статистического анализа и методов геостатистического моделирования на основе ГИС. Дж. Гидрол. Рег. Стад. 2015 , 4 , 80–110. [ Академия Google ] [ CrossRef ]
- Чаттерджи, А.; Сара, С.; Сридеви, П.Д.; Селлес, А.; Ахмед, С. Демаркация зон уязвимости фторида в гранитном водоносном горизонте, полузасушливый регион, Теленгана, Индия. Араб. Дж. Геоски. 2017 , 10 , 558. [ Google Scholar ] [ CrossRef ]
- Джампани, М.; Хюльсманн, С.; Лидл, Р.; Сонкамбл, С.; Ахмед, С.; Амерасингхе, П. Пространственно-временное распределение и химическая характеристика качества подземных вод орошаемой системы сточных вод: тематическое исследование. наук. Тотальная среда. 2018 , 636 , 1089–1098. [ Академия Google ] [ CrossRef ]
- Бро, Р. Учебное пособие по хемометрике и интеллектуальным лабораторным системам PARAFAC. Учебное пособие и приложения ; Elsevier BV: Амстердам, Нидерланды, 1997. [ Google Scholar ]
- Меллор, ФП; Сей, Э.Э. Использование обобщенных аддитивных смешанных моделей для оценки пространственного, временного и гидрологического контроля над бактериями и нитратами в уязвимом сельскохозяйственном водоносном горизонте. Дж. Контам. Гидрол. 2015 , 182 , 104–116. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Вуд, С.Н. Обобщенные аддитивные модели: введение в R ; CRC Press: Бока-Ратон, Флорида, США, 2017. [ Google Scholar ]
- Ван, С.; Тан, К.; Песня, X.; Юань, Р.; Хан, З.; Пан, Ю. Факторы, способствующие загрязнению нитратами в зоне пополнения подземных вод Северо-Китайской равнины. Гидрол. Процесс. 2016 , 30 , 2271–2285. [ Академия Google ] [ CrossRef ]
- Цао, Ю.; Тан, К.; Песня, X.; Лю, К.; Чжан Ю. Время пребывания как ключ к комплексной оценке взаимосвязи между изменением землепользования и нитратами в региональных системах подземных вод. Окружающая среда. наук. Процесс. Воздействие 2013 , 15 , 876–885. [ Академия Google ] [ CrossRef ]
- Ван, С.; Тан, К.; Песня, X.; Ван, К.; Чжан, Ю.; Юань, Р. Влияние линейного резервуара сточных вод на пополнение подземных вод и геохимическую эволюцию в полузасушливой зоне водораздела озера Байяндянь, Северо-Китайская равнина. наук. Тотальная среда. 2014 , 482–483 , 325–335. [ Академия Google ] [ CrossRef ]
- Чжан, В.; Ченг, Дж. Х.; Сиань, QS; Кюи, Дж. Ф.; Тан, XY; Ван, Г.С. Динамика и источники коллоидов в неглубоких грунтовых водах в низинных колодцах и гидроразрывном потоке на наклонных сельскохозяйственных угодьях. Вода Рес. 2019 , 156 , 252–263. [ Академия Google ] [ CrossRef ]
- Маклин, Мичиган; Эверс, Л.; Боуман, AW; Бонте, М.; Джонс, У.Р. Статистическое моделирование данных мониторинга загрязнения подземных вод: сравнение пространственных и пространственно-временных методов. наук. Тотальная среда. 2019 , 652 , 1339–1346. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Шривастава, Д.; Сингх, Р.М. Характеристика кривых прорыва и идентификация неизвестного источника загрязнения в системе подземных вод с использованием искусственной нейронной сети (ИНС). Окружающая среда. Судмедэкспертиза 2014 , 15 , 175–189. [ Академия Google ] [ CrossRef ]
- Пан, З.; Лу, В.; Чанг, З.; Ван, Х. Одновременная идентификация пространственно-временных характеристик источника загрязнения подземных вод и гидравлических параметров на основе гибридного эвристического алгоритма нейронной сети с глубокой регуляризацией. Дж. Гидрол. 2021 , 600 , 126586. [ Академика Google ] [ CrossRef ]
- Хуан, Х.; Ли, Х.; Чжоу, Дж.; Лю, В.; Ли, Дж.; Лю, Б.; Си, Б.; Цзян, Ю. Раннее предупреждение о загрязнении подземных вод на основе модели QTR для регионального управления рисками: тематическое исследование в городе Лоян, Китай. Окружающая среда. Загрязнение. 2020 , 259 , 113900. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Чаудхури, С.; Але, С. Оценка долгосрочного (1960–2010 гг.) загрязнения подземных вод фторидами в Техасе. Дж. Энвайрон. Квал. 2014 , 43 , 1404–1416. [ Академия Google ] [ CrossRef ]
- Моран, П.П. Заметки о непрерывных стохастических явлениях. Биометрика 1950 , 37 , 17–23. [ Академия Google ] [ CrossRef ]
- Чаудхури, С.; Але, С.; ДеЛон, П.; Раджан, Н. Пространственно-временная изменчивость концентрации нитратов в грунтовых водах в Техасе: с 1960 по 2010 годы. J. Environ. Qual 2012 , 41 , 1806–1817 гг. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Баши-Азгади, С.Н.; Керакян, Р.; Базарган-Лари, MR; Никоо, М.Р. Идентификация источников загрязнения в системах подземных вод: применение теории сожаления и байесовских сетей. Иран. Дж. Наук. Технол. — Пер. Гражданский. англ. 2016 , 40 , 241–249. [ Академия Google ] [ CrossRef ]
- Стамп, К.; Журек, AJ; Вахнев, П.; Гарджини, А.; Гемици, А.; Филиппини, М.; Витчак, С. Инструмент дерева решений, поддерживающий оценку уязвимости подземных вод. Окружающая среда. Науки о Земле. 2016 , 75 , 1057. [ Google Scholar ] [ CrossRef ]
- Феррейра, Д.; Алмейда, Дж.; Саймс, М.; ПресМартин, М. Сельскохозяйственная практика и геостатистическая оценка загрязнения нитратами подземных вод в районе бассейна реки Хукар, Испания. Эмир. Дж. Продовольственное сельское хозяйство. 2016 , 28 , 415. [ Google Scholar ] [ CrossRef ]
- Перес-Мартин, М.А. Распределенная имитационная модель гидрологического цикла и качества воды, интегрированная в географические информационные системы для крупных бассейнов. Вклад в анализ давления и последствий Рамочной директивы по воде ; Модель Distribuido de Simulac; Политехнический университет Валенсии: Валенсия, Испания, 2005 г. [ Google Scholar ]
- Ким, Х.; Сандерсен, ПБ; Якобсен, Р.; Каллесё, AJ; Клаас, Н.; Блихер-Матисен, Г.; Фогед, Н.; Ааманд, Дж.; Хансен, Б. Трехмерная гидрогеохимическая модель переноса и судьбы нитратов в водосборе ледниковых отложений: первый шаг к цифровой модели. наук. Тотальная среда. 2021 , 776 , 146041. [ Академика Google ] [ CrossRef ]
- Мишель эль Макдональд, Дж.; Харбо, А.В. Методы исследования водных ресурсов Объединенной геологической службы, Modu Finite-Diffe ; Публикации Геологической службы США: Денвер, Колорадо, США, 1988 г.
- Веллман, Т.П.; Кауфман, Л.; Кларк, Б. Зональная оценка внутренней восприимчивости избранных основных водоносных горизонтов Соединенных Штатов. Дж. Гидрол. 2012 , 440–441 , 36–51. [ Академия Google ] [ CrossRef ]
- Чжан, Х.; Ян, Р.; Ван, Ю.; Йе, Р. Оценка и прогноз загрязнения нитратами грунтовых вод на равнине Чэнду, юго-запад Китая, связанного с сельским хозяйством. Гидрогеол. Дж. 2019 , 27 , 785–799. [ Академия Google ] [ CrossRef ]
- Ян, Л.; Чжэн, К.; Эндрюс, CB; Ван, К. Применение структуры регионального транспортного моделирования для управления нитратным загрязнением подземных вод. Подземные воды 2021 , 59 , 292–307. [ Академия Google ] [ CrossRef ]
- Шаффер, MJ; Халворсон, А.Д. Пакет выщелачивания нитратов и экономического анализа (NLEAP): Глава 13, описание и применение модели ; Американское общество почвоведения: Мэдисон, Висконсин, США, 1991. [ Google Scholar ]
- Ли, З.; Вэнь, X.; Ху, К.; Ли, Х.; Ли, С.; Чжан, X.; Ху, Б. Региональное моделирование потенциала выщелачивания нитратов из пахотных земель севооборота озимой пшеницы и яровой кукурузы на Северо-Китайской равнине с использованием модели NLEAP-GIS. Сельское хозяйство. Экосист. Окружающая среда. 2020 , 294 , 106861. [ Академика Google ] [ CrossRef ]
- Куракос, Г.; Кляйн, Ф.; Кортис, А.; Хартер, Т. Система моделирования загрязнения из неточечных источников подземных вод для оценки долгосрочной динамики вероятностей превышения содержания загрязняющих веществ в колодцах и других местах сброса. Водный ресурс. Рез. 2012 , 48 . [ Академия Google ] [ CrossRef ]
- Банчери, М.; Коппола, А.; Базиль А. Новая модель передаточной функции для оценки времени перемещения растворенных веществ без точечных источников. Дж. Гидрол. 2021 , 598 , 126157. [ Академика Google ] [ CrossRef ]
- Ахаван, С.; Абеди-Купай, Дж.; Мусави, Сан-Франциско; Афьюни, М.; Эсламян, СС; Аббаспур, К.К. Применение модели SWAT для исследования выщелачивания нитратов в водоразделе Хамадан-Бахар, Иран. Сельское хозяйство. Экосист. Окружающая среда. 2010 , 139 , 675–688. [ Академия Google ] [ CrossRef ]
- Као, Ю.Х.; Лю, CW; Джанг, CS; Зан, Юго-Запад; Лин, К.Х. Оценка загрязнения азотом грунтовых вод на рисовых и горных полях. Пэдди-Уотер-Эквайронвайр. 2011 , 9 , 301–307. [ Академия Google ] [ CrossRef ]
- Паркхерст, Л.; Аппело, Руководство пользователя CAJ по Phreeqc (Версия 2) — компьютерной программе для видообразования, пакетного реагирования, одномерного переноса и обратных геохимических расчетов. Водный ресурс. Расследование. Rep. 1999 , 99 , 312. [ Google Scholar ]
- Лин, Дж. Х.; Ричардс, доктор медицинских наук; Ага, РГ-Т.; Ченг, младший; Ченг, Х.-П.; Джонс, Н.Л. FEMWATER: Трехмерная компьютерная модель конечных элементов для моделирования зависящего от плотности потока и транспорта в средах с переменной насыщенностью ; Экспериментальная станция инженера водных путей армии США: Виксбург, Массачусетс, США, 1997 г.
- Ллопис-Альберт, К.; Паласиос-Маркес, Д.; Мериго, Дж. М. Связанная стохастическая система обратного управления для борьбы с неточечным сельскохозяйственным загрязнением в условиях неопределенности параметров подземных вод. Дж. Гидрол. 2014 , 511 , 10–16. [ Академия Google ] [ CrossRef ]
- Пенья-Аро, С.; Пулидо-Веласкес, М.; Саукильо, А. Основа гидроэкономического моделирования для оптимального управления загрязнением подземных вод нитратами в результате сельского хозяйства. Дж. Гидрол. 2009 , 373 , 193–203. [ Академия Google ] [ CrossRef ]
- Чжан, З.; Чен, X.; Ченг, К.; Ли, С.; Юэ, Ф.; Пэн, Т.; Уолдрон, С.; Оливер, DM; Соулсби, К. Совместное гидрологическое и биогеохимическое моделирование переноса азота в карстовой критической зоне. наук. Тотальная среда. 2020 , 732 , 138902. [ Академика Google ] [ CrossRef ]
- Бейли, RT; Гейтс, ТК; Ахмади, М. Моделирование реактивного переноса селена в сочетании с азотом в орошаемой системе подземных вод регионального масштаба. Дж. Гидрол. 2014 , 515 , 29–46. [ Академия Google ] [ CrossRef ]
- Датта, Б.; Чакрабарти, Д.; Дхар, А. Идентификация неизвестных источников загрязнения подземных вод с использованием классической оптимизации со связанным моделированием. Дж. Гидро-Энвайрон. Рез. 2011 , 5 , 25–36. [ Академия Google ] [ CrossRef ]
- Восс, CI; Флорида, TAFB Имитационная модель конечных элементов для потока насыщенных и ненасыщенных, зависящих от плотности жидкости потоков подземных вод с переносом энергии или переносом химически активных однокомпонентных растворенных веществ. Водный ресурс. Расследование. Rep. 1984 , 84 , 4369. [ Google Scholar ]
- Веселинов В.В.; Александров, Б.С.; О'Мэлли, Д. Идентификация источника загрязнения с использованием полуконтролируемого машинного обучения. Дж. Контам Гидрол. 2018 , 212 , 134–142. [ Академия Google ] [ CrossRef ]
- Белушрани, А.; Абед-Мераим, К.; Кардосо, Ж.-Ф.; Мулен, Э. Метод слепого разделения источников с использованием статистики второго порядка. IEEE Транс. Сигнальный процесс. 1997 , 45 , 434–444. [ Академия Google ] [ CrossRef ]
- Ли, Д.; Сын, Х.С. Изучение частей объектов путем факторизации неотрицательной матрицы. Летт. Нат. 1999 , 401 , 788–791. [ Академия Google ] [ CrossRef ]
- Александров С.; Веселинов В.В. Слепое разделение источников для анализа давления подземных вод на основе неотрицательной матричной факторизации. Водный ресурс. Рез. 2014 , 50 , 7332–7347. [ Академия Google ] [ CrossRef ]
- Боттрелл, Ш.; Форд, Д.; Уильямс, П. Карстовая геоморфология и гидрология ; John Wiley & Sons: Хобокен, Нью-Джерси, США, 1991 г.; Том 157. [ Google Scholar ] [ CrossRef ]
- Альбукерке, штат Мэриленд; Санс, Г.; Оливейра, Сан-Франциско; Мартинес-Алегрия, Р.; Антунес, IMHR Пространственно-временная оценка уязвимости подземных вод — совмещенный подход дистанционного зондирования и ГИС для реконструкции исторического земного покрова. Водный ресурс. Менеджер. 2013 , 27 , 4509–4526. [ Академия Google ] [ CrossRef ]
- Элиас, Д.; Ангелики, М.; Василики, М.; Мария, Т.; Кристина З. Геопространственные исследования загрязнения и качества воды подземных вод, подтвержденные спутниковыми данными: пример реки Эврос (Восточное Средиземноморье). Чистое приложение. Геофиз. 2012 , 171 , 977–995. [ Академия Google ] [ CrossRef ]
- Виас, М.; Андрео, Б.; Перлз, MJ; Карраско, Ф.; Вадилло, И.; Хименес, П. Предлагаемый метод картирования уязвимости подземных вод в карбонатных (карстовых) водоносных горизонтах: метод COP. Гидрогеол. Дж. 2006 , 14 , 912–925. [ Академия Google ] [ CrossRef ]
- Тюрпо, П.; Зини, Л.; Равбар, Н.; Куччи, Ф.; Петрич, М.; Урбанк, Дж. Разработка протокола зонирования карстовых источников воды: применение к классическому карстовому региону (северо-восток Италии и юго-запад Словении). Водный ресурс. Менеджер. 2018 , 32 , 1953–1968. [ Академия Google ] [ CrossRef ]
- Живанович, В.; Жемцов И.; Драгишич, В.; Атанакович, Н.; Магазинович, С. Защита карстовых источников подземных вод на основе модели оценки уязвимости, зависящей от времени: тематическое исследование источников Црница, Восточная Сербия. Окружающая среда. Науки о Земле. 2016 , 75 , 1224. [ Google Scholar ] [ CrossRef ]
- Живанович, В.; Атанакович, Н.; Стоядинович, С. Оценка уязвимости как основа для разграничения санитарных зон карстовых источников подземных вод – практический пример источника Бледерия. Вода 2021 , 13 , 2775. [ Google Scholar ] [ CrossRef ]
- Дедевану, М.; Бине, С.; Руэ, Ж.Л.; Коке, Ю.; Брюанд, А.; Ноэль, Х. Уязвимость подземных вод и картирование рисков на основе распределения времени пребывания: пространственный анализ для оценки сосредоточенных параметров. Водный ресурс. Менеджер. 2015 , 29 , 5489–5504. [ Академия Google ] [ CrossRef ]
- Хакун, В.; Орбан, П.; Дассарг, А.; Бруйер, С. Факторы, контролирующие пространственные и временные закономерности содержания множества пестицидных соединений в подземных водах (меловой водоносный горизонт Эсбай, Бельгия). Окружающая среда. Загрязнение. 2017 , 223 , 185–199. [ Академия Google ] [ CrossRef ]
- Ромшу, ЮАР; Дар, РА; Муртаза, КО; Рашид И.; Дар, Ф.А. Гидрохимическая характеристика и оценка загрязнения подземных вод в Джамму Сиваликсе, Индия. Окружающая среда. Монит. Оценивать. 2017 , 189 , 122. [ Google Scholar ] [ CrossRef ]
- Торрес, Нью-Йорк; Ю, Х.; Падилья, И.Ю.; Маккиавелли, RE; Гасемизаде, Р.; Каэли, Д.; Кордеро, Дж. Ф.; Микер, доктор медицинских наук; Альшавабке, А.Н. Влияние гидрогеологических и антропогенных переменных на загрязнение фталатами в эогенетически-карстовых системах подземных вод. Окружающая среда. Загрязнение. 2018 , 237 , 298–307. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Ю, Х.; Гасемизаде, Р.; Падилья, И.; Иризарри, К.; Каэли, Д.; Альшавабке, А. Пространственно-временные изменения концентрации CVOC в карстовых водоносных горизонтах: анализ данных за три десятилетия из Пуэрто-Рико. наук. Тотальная среда. 2015 , 511 , 1–10. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Бутчер, К.; Хаггенбергер, П. Влияние на гидрологию карста трехмерного геологического моделирования с использованием градиентного подхода основания водоносного горизонта. Дж. Гидрол. 2007 , 342 , 184–198. [ Академия Google ] [ CrossRef ]
- Эптинг, Дж.; Пейдж, РМ; Окенталер, А.; Хаггенбергер, П. Мониторинг и моделирование карстовых источников на основе процессов — связь между внутренней и конкретной уязвимостью. наук. Тотальная среда. 2018 , 625 , 403–415. [ Академия Google ] [ CrossRef ] [ PubMed ]
- Ли, К.; Ци, Дж.; Син, З.; Ли, С.; Цзян, Ю.; Даниэлеску, С.; Чжу, Х.; Вэй, X.; Мэн, Ф.-Р. Подход к оценке воздействия землепользования и биофизических условий ландшафта на скорость пополнения и азотную нагрузку подземных вод. Сельское хозяйство. Экосист. Окружающая среда. 2014 , 196 , 114–124. [ Академия Google ] [ CrossRef ]
- Кавури, П.; Карацас, врач общей практики; Плань, В. Пара моделей южной воды и методология картографии уязвимых мест для работы с карстическими ландшафтами. Гидрогеол. Дж. 2017 , 25 , 1301–1317. [ Академия Google ] [ CrossRef ]
- Дирш, Х.-Й.Г. Теплоперенос в пористых средах. В FEFLOW ; Springer: Берлин/Гейдельберг, Германия, 2014 г.; стр. 673–709. [ Академия Google ] [ CrossRef ]
- Бине, С.; Жуано, Э.; Пауэлс, Х.; Альберик, П.; Флехок, К.; Брюанд, А. Водообмен, смешивание и временное хранение между насыщенным карстовым каналом и окружающим водоносным горизонтом: моделирование потока подземных вод и вклады стабильных изотопов воды. Дж. Гидрол. 2017 , 544 , 278–289. [ Академия Google ] [ CrossRef ]
- Сурдык, Н.; Гутьеррес, А.; Баран, Н.; Тьери, Д. Сосредоточенная модель для моделирования изменения концентрации нитратов в подземных водах в масштабе водосбора. Дж. Гидрол. 2021 , 596 , 125696. [ Академика Google ] [ CrossRef ]
- Тьери, Д.; Сеген, Ж.-Ж. Глобальное моделирование переноса нитратов в гидрогеологическом бассейне для предыдущего этапа L'évolution Des Concentrations в Les Eaux Souterraines Описание Du Modele BICHE Trois Exemples D'application ; Bureau De Recherches Géologiques Et Minières Service Géologique National Département Eau: Орлеан, Франция, 1985.
Страница 8 из 30